脂質は膵臓のリパーゼで消化されて,脂肪酸とグリセリンになる。グリセリンはグリセロール-3-リン酸を経てジヒドロキシアセトンリン酸 (DHAP)
になり,解糖経路に入って代謝される(トリグリセリドの合成の項を参照)。一方,脂肪酸はミトコンドリアに運ばれた後,b-酸化によってアセチル-CoAにまで代謝される。
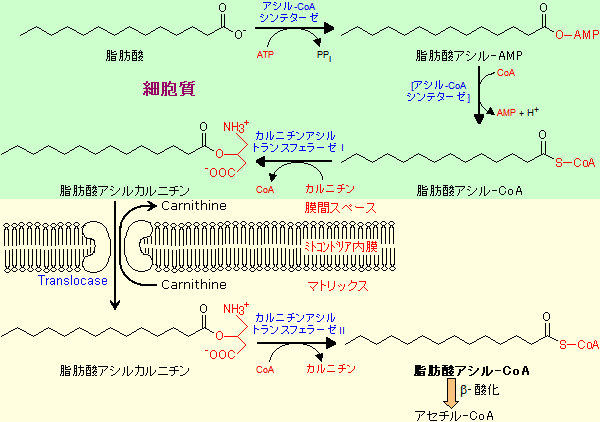
長鎖脂肪酸は細胞質で活性化されてアシル-CoAになるが、アシル-CoAはミトコンドリア内膜をを通過できない。そのため、一端、カルニチンと結合してからミトコンドリアのマトリックス内に取り込まれ、再び脂肪酸アシル-CoAに再生される。カルニチン[carnithine]やアシル-カルニチンはミトコンドリア内膜を通る事ができる。
ミトコンドリアに取り込まれた脂肪酸アシル-CoAはb‐酸化の出発原料になり,アセチル-CoAにまで分解される。
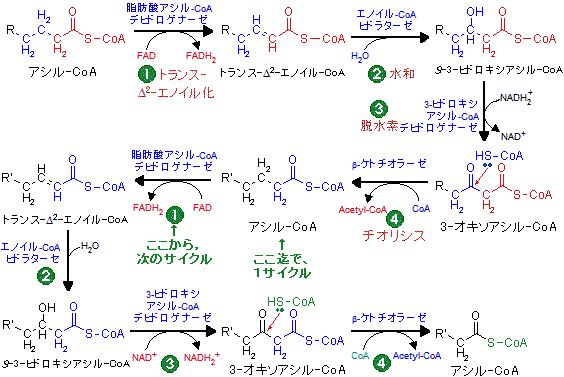
偶数炭素数の長鎖脂肪酸は、上に示したように脂肪酸アシル-CoAになってミトコンドリアのマトリックスに取り込まれる。この脂肪酸アシル-CoAを出発原料として、以下のサイクル様の代謝経路によって、最終的にアセチル-CoAになる。この経路をb‐酸化という。β-酸化は、 トランス-D2-エノイル化、
トランス-D2-エノイル化、 水和、
水和、 脱水素化、
脱水素化、 チオリシスの4つの段階から成る。
チオリシスの4つの段階から成る。
脂肪酸のC-C結合を切断するには,b位を活性メチレンに変える戦略が採られる。反応で得られる3-オキソアシル-CoAはCH2が2つのカルボニル基で挟まれているので,カルボニル基の電子吸引性のために特にβ位で切れ易い構造となる。このようなやり方は,六炭糖のC3-C4間を切るために,グルコースをフルクトースに異性化する場合にも見られる。
次にアシル-CoAからアセチル-CoAに到る経路を示す。
|
 |
| アシル-CoAが全てアセチル-CoAになる迄、b-酸化が続く |
 |
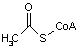
アセチル-CoA |
 脂肪酸酸化によるATPの生成量
脂肪酸酸化によるATPの生成量
- 飽和脂肪酸としてステアリン酸 (C18)を例にとる。C18から考えて,アセチル-CoAが9モルつくられる(b-酸化のサイクル数は8回)。
- ステアリン酸 → ステアロイル-CoAの変換で ATP(→AMP)を消費。これは-2 ATPに相当
9 アセチル-CoA → 9 × 12 = 108 ATPを生成(TCA回路→呼吸鎖)
8 FADH2 → 8 × 2 = 16 ATPを生成(呼吸鎖)
8 NADH2+ → 8 × 3 = 24 ATPを生成(呼吸鎖)
- よって,全体の反応は次のようになる。
- C17H35COOH + 26 O2 + 146 Pi + 146 ADP → 18 CO2 + 164 H2O + 146 ATP
- 不飽和脂肪酸の場合,(1)の段階が不要となるため(不飽和脂肪酸のb-酸化を参照)FADH2の生成がなく,その分を上のような計算から除外すれば良い。P/O比を2.5,1.5とすれば146ではなくて120 ATPとなる。
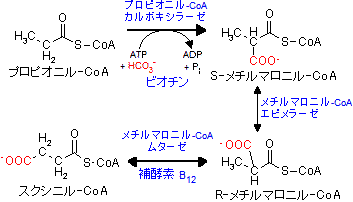
奇数炭素数の長鎖脂肪酸の場合も,偶数炭素の場合と同様にしてb-酸化が進行してアセチル-CoAとプロピオニル-CoA[Propionyl-CoA]が得られる。
最後に生じるプロピオニル-CoAは炭素数が奇数であるため,次のような過程でスクシニル-CoAにされ,TCA回路に回される。
 |
|
TCA回路へ |
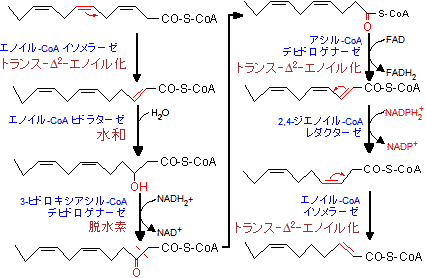
不飽和の脂肪酸の場合、二重結合の2つ手前の炭素までは通常のb-酸化が進行する。二重結合がβ位に来た所で、下の最初の反応のように、二重結合の位置がカルボニル側にずれると同時に、cis→transの変換が起こる(b-酸化では全てトランス-D2-エノイルにする必要があるから)。従って、acyl-CoA hehydrogenaseの段階がスキップされるため、FADH2の生成はない。次いで、通常の水和、脱水素、チオリシスの各反応により、アセチル-CoAが切り出される。
一方、二重結合がγ位に来た場合は、右の例のように、一旦、トランス-D2-エノイル化した後、NADPH2を補酵素として二重結合の位置が1つカルボニル側にずれ、再び、トランス-D2-エノイル化して左の最初と同じような反応経路をたどる。