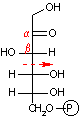
- 解糖(glycolysis)はほとんど全ての生物に共通に存在する糖の代謝経路で,反応は細胞質で行われる。解糖は Embden-Meyerhof 経路とも呼ばれ,本来、D-グルコースの嫌気的分解による乳酸やエタノール生成までの過程(発酵という)を意味したが、好気的条件下でもピルビン酸までは全く同じ経路をたどる事が分かった。ただし,ある種の微生物はEmbden-Meyerhof
経路とは少し異なるEntner-Doudoroff経路を利用する。
狭義の解糖は以下の10段階の反応で構成される。
-
解糖と糖新生の関係 赤矢印は不可逆
| グルコース(Glc) |
|
  1 1 |
|
| グルコース 6-リン酸(G-6-P) |
ピルビン酸(Pyr) |
 2 2 |
10 |
| フルクトース 6-リン酸(F-6-P) |
ホスホエノールピルビン酸(PEP) |
| 3 |
9 |
| フルクトース 1,6-ビスリン酸(FBP) |
2-ホスホグリセリン酸(2-PG) |
| 4 |
8 |
| ジヒドロキシアセトンリン酸(DHAP) |
 5 5 |
3-ホスホグリセリン酸(3-PG) |
| + |
7 |
| グリセルアルデヒド 3-リン酸(GAP) |
 |
1,3-ビスホスホグリセリン酸(BPG) |
|
6 |
|
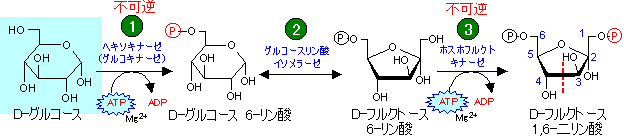
- 細胞に取り込まれたグルコースはすぐに6-リン酸化される。ATP消費。
- グルコース 6-リン酸(G-6-P)はフルクトース 6-リン酸(F-6-P)に異性化される。
(グルコースをフルクトースに異性化する理由を参照。)
- F-6-Pはさらに二リン酸に変えられる。ATP消費。
- 生じたフルクトース 1,6-ビスリン酸(FBP)はアルドール開裂により,ジヒドロキシアセトンリン酸(DHAP)とグリセルアルデヒド 3-リン酸(GAP)になる。
- DHAPはGAPに異性化される。4で分かれた2つの経路がこれで1つになる。
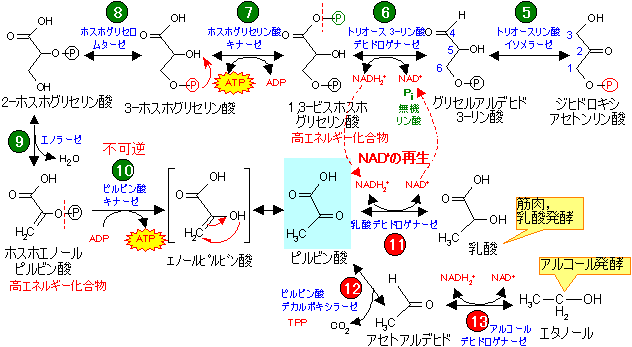
- 無機リン酸を利用してGAPを1,3-ビスホスホグリセリン酸(BPG)に変える。この段階はNAD+を消費する。
- BPGの1位のリン酸基をADPに転移し,ATPを合成(基質レベルのリン酸化)するとともに,3-ホスホグリセリン酸(3-PG)に変える。
- 3-PGを異性化する。
- 生じた2-ホスホグリセリン酸(2-PG)から,脱水反応により高エネルギー化合物であるホスホエノールピルビン酸(PEP)をつくる。
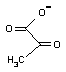
- PEPの2位のリン酸基をADPに転移し,ATPを合成(基質レベルのリン酸化)する。エノール型のピルビン酸は互変異性によりひとりでにピルビン酸になる。
|
  |
- 嫌気的条件では、乳酸やエタノールの生成が最終段階となり,1分子のグルコースから2分子のATPがつくられる。
- Glucose + 2 ADP + 2 Pi + 2 NAD+ → 2 Pyruvate + 2 ATP + 2 NADH2+ + 2 H2O
- 筋肉など大部分の組織はグリコーゲンを貯蔵しているので,解糖はグリコーゲンから始まる。この場合,グリコーゲンの分解で得られるグルコース6-リン酸(G-6-P)から出発するのでグルコース単位当たり3分子のATPがつくられる。
- G-6-P + 3 ADP + 2 Pi + 2 NAD+ → 2 Pyruvate + 3 ATP + 2 NADH2+ + 2 H2O
- 一方,好気的条件では乳酸生成の速度が著しく低下する。これは、(a) ピルビン酸→ 乳酸の経路から,(b) ピルビン酸 → アセチル-CoA → TCA回路 → 呼吸鎖の経路に切り替わるためである(パスツール効果)。(b)の経路を利用した場合,グルコース 1分子から最大38分子ものATPが得られる(グルコースの完全酸化を参照)。
しかしながら,解糖によるATP合成は呼吸鎖を利用した場合の約100倍の速度をもつため,激しい短期間の筋肉の運動などではクレアチンリン酸,次いで解糖が利用され,筋肉中に乳酸が蓄積する。生じた乳酸は肝臓に送られてピルビン酸に戻され,糖新生やTCA回路で有効に利用される(コリ回路)。
解糖の代謝流量は、その生産物に対する需要で細かく制御される。そのため,ほとんどの反応は可逆的であるが,キナーゼが触媒する反応のうち は可逆的であるが,
は可逆的であるが,

 は細胞内の条件では不可逆である(下の表でDGが大きな負の値を持つことからわかる)。従って、糖新生は単なる解糖の逆経路ではなく、不可逆的な段階では別の反応や経路を利用する事になる[「異化と同化は別経路」の1つの例]。
は細胞内の条件では不可逆である(下の表でDGが大きな負の値を持つことからわかる)。従って、糖新生は単なる解糖の逆経路ではなく、不可逆的な段階では別の反応や経路を利用する事になる[「異化と同化は別経路」の1つの例]。
解糖は以下の10段階の代謝反応より構成される。反応とはATPを消費する段階,反応とはATPを生産する段階(基質レベルのリン酸化)である。また,反応 で無機リン酸が使われ,NAD+が消費されることに注意しよう。
で無機リン酸が使われ,NAD+が消費されることに注意しよう。
|
 
|
下の図の反応番号〜 をクリックすると,より詳細な説明や酵素タンパク質の性質が見れます。 をクリックすると,より詳細な説明や酵素タンパク質の性質が見れます。 |
|
標準自由エネルギー変化(DG0')と生理的条件での
自由エネルギー変化(DG) 単位,kJ/mol
| 反応 |
酵 素 |
DG0' |
DG |
反応 |
| 1 |
ヘキソキナーゼ |
-20.9 |
-27.2 |
グルコース グルコース 6-リン酸 グルコース 6-リン酸 |
| 2 |
グルコースリン酸イソメラーゼ |
+2.2 |
-1.4 |
フルクトース 6-リン酸 |
| 3 |
ホスホフルクトキナーゼ |
-17.2 |
-25.9 |
フルクトース 1,6-ビスリン酸 |
| 4 |
アルドラーゼ |
+22.8 |
-5.9 |
ジヒドロキシアセトンリン酸
+グリセルアルデヒド 3-リン酸 |
| 5 |
トリオースリン酸イソメラーゼ |
+7.9 |
+4.4 |
ジヒドロキシアセトンリン酸
グリセルアルデヒド 3-リン酸 |
| 6 |
グリセルアルデヒド-3-リン酸デヒドロゲナーゼ |
-16.7 |
-1.1 |
1,3-ビスホスホグリセリン酸 |
| 7 |
ホスホグリセリン酸キナーゼ |
3-ホスホグリセリン酸 |
| 8 |
ホスホグリセロムターゼ |
+4.7 |
-0.6 |
2-ホスホグリセリン酸 |
| 9 |
エノラーゼ |
-3.2 |
-2.4 |
ホスホエノールピルビン酸 |
| 10 |
ピルビン酸キナーゼ |
-23.0 |
-13.9 |
ピルビン酸 |
E.A.Newsholme, C.Start, "Regulation in Metabolism",p.97, Wilew
(1973)
グルコースは解糖で2 ATPを生成するが,好気的条件下では,生じるピルビン酸を更にミトコンドリアのTCA回路を経て呼吸鎖に回すことによって,二酸化炭素と水にまで完全代謝される。この場合,解糖だけよりもっと多くのATPを生産することができる。
- グルコース → 2 ピルビン酸 + 2 ATP + 2 NADH2+ (解糖)
- 2 ピルビン酸 → 2 アセチル-CoA + 2 CO2 + 2 NADH2 (CoA化)
- 2 アセチル-CoA → 4 CO2 + 6 NADH2+ + 2 FADH2 + 2 GTP (TCA回路)
- FADH2 → 2 ATP (呼吸鎖)
 ミトコンドリアへのNADH2+の輸送
ミトコンドリアへのNADH2+の輸送
このようにして生じたNADH2+はミトコンドリアの内膜を通過できないため,2つの仕組みで膜内に輸送される。
[リンゴ酸-アスパラギン酸シャトル]肝臓,心臓,腎臓など
リンゴ酸-アスパラギン酸シャトルは,次のような輸送の仕組みにより,NADH2+をミトコンドリアにを送り込む。(NADH2+そのものは移動せず,結局,電子が移動することになる)。この輸送系を利用した場合, NADH2+から3 ATP (または2.5 ATP)がつくられる。(a-KG:a-ケトグルタル酸)
 |
[グリセロリン酸シャトル]筋肉など肝臓,心臓,腎臓以外の臓器
もう1つの輸送機構はグリセロリン酸シャトルで,ミトコンドリア内膜に結合した酵素のFAD/FADH2変換を利用して電子を送り込む。このシャトル系を利用した場合,FADH2がNADH2+の代わりとなるため,NADH2+=2 ATP (または1.5 ATP)となる。
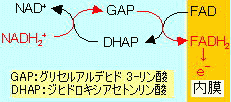 |
以上を総計すると,グルコース 1分子から,肝臓,心臓,腎臓ではNADH2+がリンゴ酸-アスパラギン酸シャトルを通過するため 38 ATP (または32 ATP),それ以外の臓器ではグリセロリン酸シャトルを通過するため 36 ATP (または30 ATP)がつくられることになる。( )内の数値はP/O比を2.5, 1.5で計算したもの。
なお,ミトコンドリアを持たない原核細胞の場合,グルコースから得られる2 NADH2+は細胞膜に存在する呼吸鎖酵素によって6 ATPに変換される。
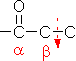
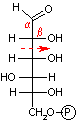
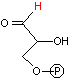
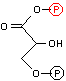
解糖の初期段階でグルコースはフルクトースに異性化される。異化代謝においてC-C結合を切断するのにしばしば用いられるやり方は,下の左図のようにカルボニル基のβ位で切ることである(脂肪酸のβ-酸化もその例)。これはカルボニル基の電子吸引性のためβメチレンが活性化されているためである。高校で習うヨードホルム反応(ハロホルム分解)はその一例である。
グルコース-6-リン酸の場合,上の図のようにC-C結合をβ位で切断するとC2とC4の化合物が生じる。一方,フルクトース-6-リン酸であればβ位が1つずれるために,2分子のC3化合物が生じる。この一方を異性化して同じ分子(グリセルアルデヒド-3-リン酸)にすることによって,以後,代謝経路を一本化できる。また,フルクトースの代謝も,フルクトース-6-リン酸という入口ができることによって,単純化できる利点もある。
嫌気的条件下では,解糖はピルビン酸を経て乳酸やエタノールの生成をもって終了する(発酵)。解糖の段階でNAD+が消費されるが,その細胞質内濃度は限られているため解糖を続行するためには再生する必要がある。そこで,経路をピルビン酸で止めずに乳酸やエタノールをつくることによっててNAD+の再生を行う必要がある。
生じた乳酸は肝臓に送られ,また元のピルビン酸に戻されて,糖新生やTCA回路で有効に利用される(コリ回路)。
 |
 |
 |
| GAP |
NAD+ NADH2+ |
1,3-BPG |
 |

 |
 |
| 乳酸 |
| 乳酸 |
デヒドロゲナーゼ |
ピルビン酸 |
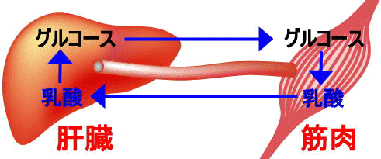
筋肉において,嫌気的条件下でグルコースから生成した乳酸は血液中に放出され肝臓へと運ばれる。肝臓で乳酸は糖新生によって再びグルコースへと変えられて血液中に放出され,筋肉に戻る。このようなグルコースと乳酸の循環をコリ回路 (Cori cycle) という。
乳酸が過剰に筋肉に蓄積すると組織のpHを低下させるため,いわゆる「疲れ」や「こり」といった現象を引き起こす。また,血液中の乳酸濃度が高くなると血液の緩衝力を超え,pHが低下する(乳酸アシドーシス)。コリ回路はそれらを解消するための肝臓と筋肉の連携による生理的な機構である。
ピルビン酸は種々の酵素によって様々な化合物に変化する。以下の例を見れば,酵素と補酵素の組み合わせによる生体内物質変化の多様性を認識できるであろう。
[代謝経路]
酢酸発酵 |
ピルビン酸 |
ピルビン酸
オキシダーゼ
 |
酢酸 + CO2 + H2O |
| H2O2 (FAD) |
| 糖新生 |
ピルビン酸 |
ピルビン酸
カルボキシラーゼ
|
オキサロ酢酸 |
| CO2 |
アルコールー
発酵 |
ピルビン酸 |
ピルビン酸デカル
ボキシラーゼ

(TPP)
|
アセトアルデヒド + CO2 |
| TCA回路 |
ピルビン酸 |
ピルビン酸
デヒドロゲナーゼ
|
アセチル-CoA + CO2 |
| CoA (TPP) |
| 乳酸発酵 |
ピルビン酸 |
乳酸
デヒドロゲナーゼ

(NAD) |
乳酸 |
アミノ酸
代謝 |
ピルビン酸 |
アラニンアミノ
トランスフェラーゼ
|
アラニン + a-ケトグルタル酸 |
| Glu (PALP) |
アミノ酸
代謝 |
ピルビン酸 |
アラニン
デヒドロゲナーゼ
|
アラニン + H2O |
| NH3 (NAD) |
グルコース以外の糖は,いずれも解糖の経路に乗せられて代謝される。
- フルクトースの代謝
| フルクトース |
フルクトキナーゼ |
フルクトース 1-リン酸 |
フルクトース 1-リン酸アルドラーゼ |
|
 |
DHAP |
 解糖へ 解糖へ |
|
ATP ADP |
|
グリセルアルデヒド |
ATP
トリオースキナーゼ
ADP |
 |
|
| グリセルアルデヒド 3-リン酸解糖へ |
- ガラクトースの代謝
| ガラクトース |
ガラクトキナーゼ |
ガラクトース 1-リン酸 |
ガラクトース 1-リン酸
ウリジルトランスフェラーゼ |
グルコース 1-リン酸 |
ホスホグルコ
ムターゼ |
グルコース 6-リン酸
 |
|
|
 |
|
ATP ADP |
UDP-グルコースUDP-ガラクトース |
|
解糖へ |
|
UDP-グルコースイソメラーゼ |
|
- マンノースの代謝
| マンノース |
ヘキソキナーゼ |
マンノース 6-リン酸 |
ホスホマンノース
イソメラーゼ |
フルクトース 6-リン酸 |
解糖へ |
|
|
|
ATP ADP |
|
エントナー・ドウドロフ経路
解糖はほとんど全ての生物に存在する糖の代謝経路(Embden-Meyerhof 経路)であるが,ある種の微生物はこれとは少し異なるエントナー・ドウドロフ (Entner-Doudoroff) 経路を利用する。
[エントナー・ドウドロフ経路] 赤矢印は解糖との違い
| グルコース 6-リン酸(G-6-P) |
|
|
|
| 6-ホスホグルコン酸 |
|
|
|
2-デヒドロ-3-デオキシ-6-ホスホグルコン酸 |
ピルビン酸(Pyr) |
|
|
| グリセルアルデヒド 3-リン酸(GAP) |
解糖 |
|
|
| ジヒドロキシアセトンリン酸(DHAP) |
1,3-ビスホスホグリセリン酸(BPG) |
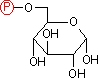
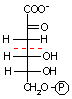
グルコース 6-リン酸 |
|
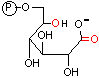
6-ホスホグルコン酸 |
ホスホグルコン酸
デヒドラターゼ |
 |
|
 |
HMSの
と同様 |
H2O |
|
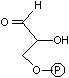
2-デヒドロ-3-デオキシ-
6-ホスホグルコン酸 |
 |
2-デヒドロ-3-デオキシ-
6-ホスホグルコン酸
アルドラーゼ |
|
エタノール,乳酸← |
|
 |
|
ピルビン酸 |
グリセルアルデヒド
3-リン酸 |
 |
解糖経路 |
| 2ATP NADH2+ |
NAD+ 2ADP |
- ピロ解糖系
古細菌では通常の解糖系(Embden-Meyerhof 経路)は働かず,エントナー・ドウドロフ経路と下記のピロ解糖系を利用する。
-
[ピロ解糖系] 赤矢印は解糖との違い
| グルコース |
|
|
|
| グルコン酸 |
|
|
| 2-デヒドロ-3-デオキシグルコン酸 |
ピルビン酸(Pyr)PEP |
|
グリセルアルデヒドDH |
グリセリン酸キナーゼ |
| グリセルアルデヒドグリセリン |
酸2-ホスホグリセリン酸 |
|
アルドースレダクターゼ |
|
| グリセロール |
3--ホスホグリセリン酸 |
|
グリセロールキナーゼ |
|
| グリセロール 3-リン酸 |
1,3-ビスホスホグリセリン酸 |
|
|
|
| ジヒドロキシアセトンリン酸(DHAP) |
グリセルアルデヒド 3-リン酸 |