|
�y�f�̕����@�y�f�����̓����@�y�f�������x�_�@�y�f�����̒��� |
| �y�f�����̈�ʓI�ȕ\���� | |
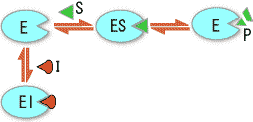
| E + S �@E�F�y�f�@S�F��@ES�F�y�f-������́@P�F������ 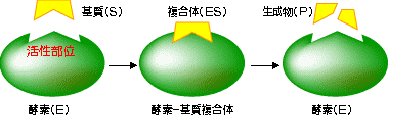 �@ �@ |
|
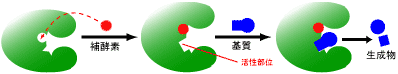
| �y�f�����͍y�f�Ɗ�������̂����邱�Ƃ���n�܂�B | |
 �y�f�����̗��j
�y�f�����̗��j
|
|
  |
| �y�f | �����̌`�� | �� |
|---|---|---|
| 1. �_���Ҍ��y�f (Oxidoreductase) |
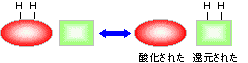
�_���Ҍ����� | �f�q�h���Q�i�[�[�Q �m2���f���p�n�C�V�g�N�����Q �mFe2+/3+���p�n�C �J�^���[�[�C�I�L�V�_�[�[�Q�C �I�L�V�Q�i�[�[�Q�C���b�_�s�O�a���y�f |
 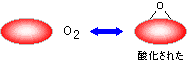 |
||
| 2. �]�ڍy�f (Transferase) |
���q�c�]�ڔ��� | �A�V���]�ڍy�f �m�A�V����]�ځn�C�L�i�[�[�Q �m�����_��]�ځn�C �A�~�m�g�����X�t�F���[�[�Q �m�A�~�m��]�ځn |
 |
||
| 3. ���������y�f (Hydrolase) |
�������� | �^���p�N�������y�f�Q �i�v���e�A�[�[�j�C ���������y�f�Q�i���p�[�[�j�C ���������y�f�Q�i�A�~���[�[�C���]�`�[���C���]�K���N�g�V�_�[�[�j �����_�����y�f�Q�i�k�N���A�[�[�Q�C�z�X�t�@�^�[�[�Q�C�����y�f�j ���̑��i�E���A�[�[�CATP���������y�f�j |
 |
||
| 4. �E���y�f (Lyase) |
�t������ђE������ | �Y�_�q�h���^�[�[�C�s���r���_�f�J���{�L�V���[�[ |
 |
||
| 5. �ِ����y�f (Isomerase) |
�ِ������� | ���Z�}�[�[�Q�C�z�X�z�O���Z�����_�z�X�z���^�[�[�C �O���R�[�X6-�����_�C�\�����[�[ |
 |
||
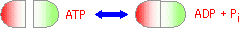
| 6. �����y�f (Ligase, Synthetase) |
C-C, C-O, C-N���� �Ȃǂ̐������� �iATP��v���j |
DNA���K�[�[�C�A�~�m�A�V����RNA�����y�f�C �A�V��CoA�V���e�^�[�[�C�J���{�L�V���[�[�Q |
 |
||
�⏕���q:
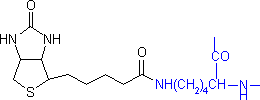 �r�I�`�j����iLys�c��Ɍ����j
�r�I�`�j����iLys�c��Ɍ����j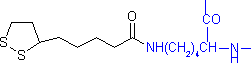 ���|�C����iLys�c��Ɍ����j
���|�C����iLys�c��Ɍ����j
 ���̓A�|�y�f�i�s�����j��\���B�⏕���q(��)���A�|�y�f�Ɍ�������Ɗ����^�̃z���y�f�ɂȂ�C��ɍ�p�ł���B |
|
|
|
|
�y�f�Ɗ������G�l���M�[
\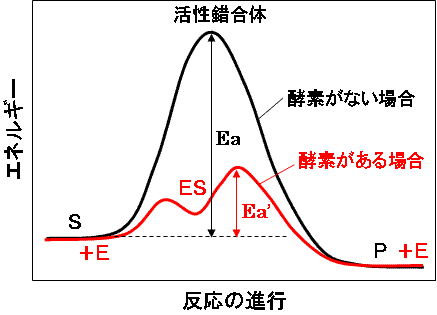 |
Ea,Ea'�F�������G�l���M�[ |
|||||||||
| [�������G�l���M�[�ƐG�}] |
| Arrhenius�̎� | |
| log k = A/2.303 - Ea/2.303RT | |
| k, �������x�萔�G �`�C�p�x���q�G �q�C�C�̒萔�i��8.31 J/mol�EK�j�G �s�C��Ή��x | |
| �ߎ_�����f�̕����F�@�����G�}�ƍy�f�J�^���[�[�̐G�}�\�̔�r | |
| log(kcat/kPt) = (5 X 104 - 2 X 104)/(2.303 X 8.31 X 310) = 5.06 | |
| �� kcat/kPt ��105 �J�^���[�[�͔������10���{���G�}�\�������I���G�}�ɔ�ׂ�C�����{�ɂȂ�B |
|
�y�f�̐G�}��p�̓���
 ���K���x(optimum temperature)
���K���x(optimum temperature)
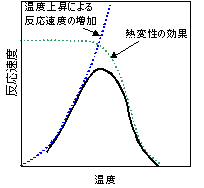 |
| [�y�f�������x�Ɖ��x] |
���KpH (optimum pH)
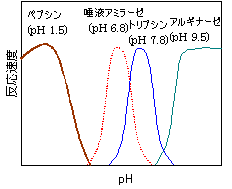 |
| [�y�f�������x��pH] |
����ِ�(substrate specificity)
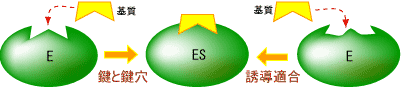
|
|
|
|
�~�J�G���X�E�����e���̎�
| ��1 | ��cat | |||||
| �d �{ �r | �d�r | �d �{ �o | �i�P�j | |||
| ��2 | ||||||
| �d�r�̐������x�́A | ||||||
| ��[�d�r�n
���� |
����1�m�d][�r�n�|�i��2�{��cat�j[�d�r�n | �i�Q�j | ||||
| �y�f����ъ�̏��Z�x���m�d�n0�A�m�r�n0�Ƃ���ƁA | ||||||
| �m�d�n0���m�d�n�{�m�d�r�n | �i�R�j | |||||
| �m�r�n0���m�r�n�{�m�d�r�n�{�m�o�n | �i�S�j | |||||
| �ʏ�̎��������ł́m�r�n0��m�d�n0�ł���A�����̏������Ƃ�C�������𗦂�10���ȉ��̏ꍇ�́C�m�o�n�͖����ł��邩��A | ||||||
| �m�r�n0���m�r�n | �i�T�j | |||||
| �Ƃ��Ă悢�B �܂��A�����̂��������ȊO�́A�m�d�r�n�̔Z�x�ω��͂Ȃ��ƍl���ėǂ��i����ԋߎ��j�̂ŁA�i�Q�j���A�i�R�j���A�i�T�j�����A |
||||||
| ��[�d�r]
���� |
����1([�d]0�|[�d�r])[�r]0�|(��2�{��cat)[�d�r]���O�@�@�i�U�j | |||||
| ������܂Ƃ߂�ƁA | ||||||
| [�d�r]�� | ��1�m�d]0�m�r]0 ��1�m�r]0�{��2�{��cat �@ |
|||||
| �� | �@�@�@�m�d]0�m�r]0 �m�r]0�{(��2�{��cat)/��1 |
�i�V�j | ||||
| �������x �� �́A | ||||||
| �� �� ��cat�m�d�r�n | �i�W�j | |||||
| ������i�V�j���ɑ�����āA | ||||||
| �� �� | ��cat�m�d]0�m�r]0
�m�r]0�{�jm |
�i�X�j | ||||
| ������ �jm �̓~�J�G���X�萔�ŁA�����ŗ^������B | ||||||
| �jm �� | ��2 �{��cat
��1 |
�i10�j | ||||
| ���܁A�m�r�n��jm �̏������l����ƁA�i�X�j�����A | ||||||
| �� �� | ��cat�m�d]0 | �i11�j | ||||
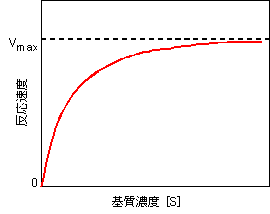
| �ƂȂ�A�������x�́m�r]0 �Ɉˑ������O�������ƂȂ�B ���̎��� �� �� �umax �Ƃ���ƁA |
||||||
| �umax�� | ��cat�m�d]0 | �i12�j | ||||
| �ƂȂ�B�y�f�Z�x�����̏����ő��肷��A�umax�͒萔�ƂȂ�B �]���āA�i�X�j���͎��̂悤�ɏ��������ł���B |
||||||
| �� �� | �umax�m�r]0
�m�r]0�{�jm |
�i�~�J�G���X�E�����e���̎��j �i13�j | ||||
| �@��Z�x�Ɣ������x�̊W��}�Ŏ����ƁC���̂悤�ɂȂ�B | ||||||
 |
||||||
�~�J�G���X�萔�̈Ӗ�
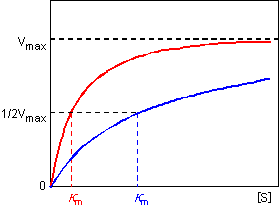 ������ɑ��āC�jm ���قȂ�y�f�̏ꍇ�C�jm ���������قǍ�p�������Ƃ����顓��l�ɁC�����y�f�ɑ��āC�jm ���قȂ��̏ꍇ�C�jm ����������قǍ�p���₷���Ƃ����� |
||||||||||||||
�y�f-������́i�d�r�j�̉𗣒萔 �jS �͎��̂悤�ɗ^������B
�@�]���āA�jm �l���������� �d�r�� �𗣂��N���ɂ����A�܂�A�y�f�Ɗ���������Ղ����ƂɂȂ�B ��ʂɁAE + S �� ES �̔����͋ɂ߂đ������t�ɂ���A��1�A��2����cat�̏�������������Ă��邱�Ƃ��������Ă���B����������t�̎�舵���ƌĂԁB |
||||||||||||||
| �m�r�n��jm�̏ꍇ�i��Z�x�̊�j�F |
|||||
| �~�J�G���X�E�����e���̎����A | |||||
| �� �� | �i�umax�^�jm�j�m�r]0 | �i15�j | |||
| �ƂȂ�A�����͊�Z�x�ɔ�Ⴕ�C�ꎟ�����ƂȂ�B | |||||
| �jm��m�r�n�̏ꍇ�i���Z�x�̊�j�F |
|||||
| �� �� | �umax | �i16�j | |||
| �Ȃ̂ŁC�O�������ɂȂ�C�������x�͊�Z�x�ɖ��W�ł��� | |||||
�~�J�G���X�萔�̋��ߕ�
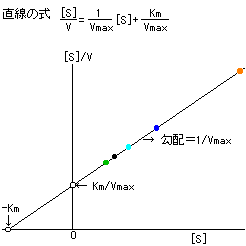
 Lineweaver-Burk�v���b�g Lineweaver-Burk�v���b�g |
|
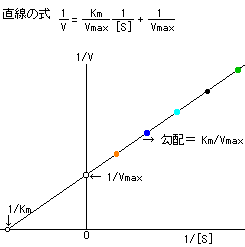 |
Lineweaver-Burk�v���b�g�ł́A������1/[S]�A�c���1/V���Ƃ�܂��B������X�ؕЂ� -1/Km�AY�ؕЂ�1/Vmax�ƂȂ�̂ŁA�O���t����Km�AVmax�����߂邱�Ƃ��o���܂��B Lineweaver-Burk�v���b�g�́A�Ⴂ��Z�x�̃f�[�^�̌덷�̉e�����₷���Ƃ������_������A3��̃v���b�g�̒��ł͍ł������v���b�g�ƌ����܂��B�]���āA���̃v���b�g�@�łقڒ����ɂȂ�������Ƃ����ăf�[�^�̎����ǂ����ǂ����͕�����܂���B |
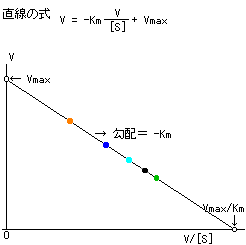
| Eadie-Hofstee�v���b�g |
Hanes-Woolf�v���b�g |
 |
 |
| ��̐}�Ɏ����悤�ɁAv/[S]�����̒l�Ńv���b�g����̂��AEadie-Hofstee�v���b�g�ł��B���̃v���b�g�͔������x�̌덷�ɕq���Ȃ̂������ł��i�܂�A�����Ƃ��V�r�A�ȕ��@�ƌ�����j�B�f�[�^�̐��x���ǂ��ꍇ�́C������Michaelis-Menten�̎��ɍ��v���邩�ǂ����������o���̂ɗL�p�ł��B | Hanes-Woolf�v���b�g�͊�Z�x [S]��[S]/���Ńv���b�g���܂��B�������x�̌덷��Km�� Vmax�̌���ɋy�ڂ��e���𑍍��I�ɍl������ƁA���̃v���b�g�@���ł��Ó��ȕ��@�Ƃ����Ă��܂��B |
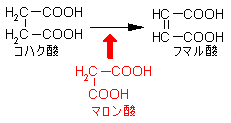
�y�f�����̑j�Q
|
|||||||||||||||||
�h�R�j�Q�܂̈��
|
|||||||||||||||
��h�R�j�Q��
| �E�y�f�̂������C�I���ƍ������`��������́F�@CN-, H2S, CO | |
| �J�^���[�[�C�y�[�I�L�V�_�[�[�C�V�g�N�����ށcFe �|���t�F�m�[���I�L�V�_�[�[�cCu �J���{�j�b�N�A���q�h���[�[�cZn |
|
| �E�d�����C�I���CHg, Ag�Ȃ� | |
| �E���A�[�[�C�p�p�C�� | |
| �E�_���� | |
| SH�y�f��SH���-S-S-�Ɏ_�����ĕs������ | |
| �E�E�ʊ����� | |
| �y�f�̑a�����\�ʂ⊈�����ʋߕӂɋz�����ĕs������ | |
| �s�h�R�j�Q�̑��x�_ |
 |
|
 |
||
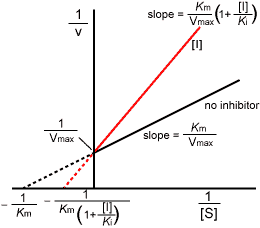
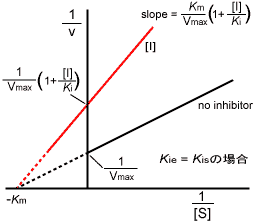
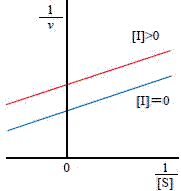
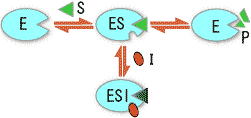
| �~�J�G���X�����e���̎��͎��̂悤�ɂȂ�B | ||
| v �� �umax�m�r]/( ���m�r]�{�jm) | ||
| �@�������C����1 + [I]/�j�� | ||
| Lineweaver-Burk�v���b�g�̎��͎��̂悤�ɂȂ�B | ||
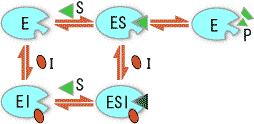
������������j�Q��(tight binding inhibitors)
|
|
|
|
�s�����ȑO��^���p�N���Ƃ��Ă����C�y�v�`�h���̈ꕔ�����Ċ����^�̍y�f�ɕω�������̂�����B
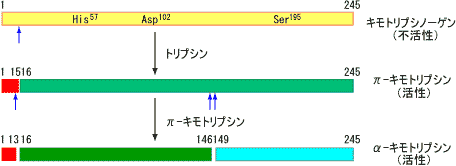 �L���g���v�V�m�[�Q���̊������@�\ |
|
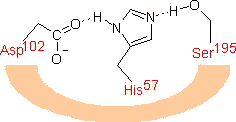 �Z�����v���e�A�[�[�� �������S�c�� |
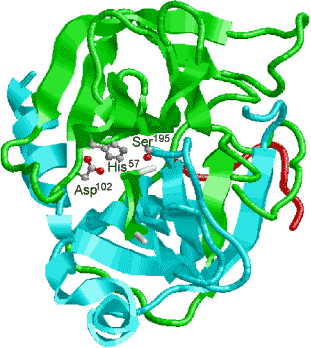 ��-�L���g���v�V���̗��̍\�� |
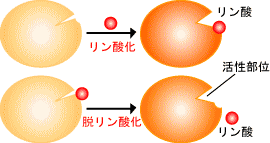
�����_���ƒE�����_���ɂ��y�f�����̒���
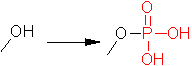 |
 |
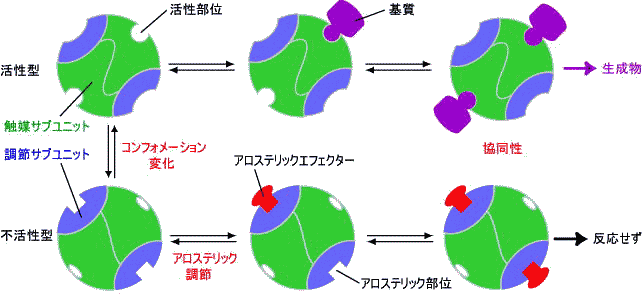
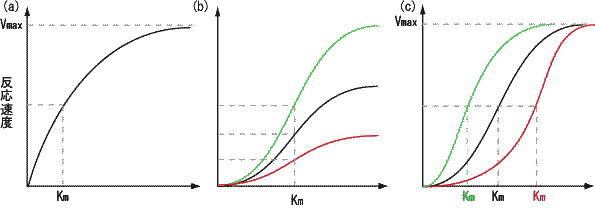
�A���X�e���b�N�y�f
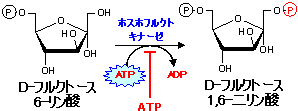
 �t�B�[�h�o�b�N�j�Q�̗�
�t�B�[�h�o�b�N�j�Q�̗�
|
|