
|
ATP NAD+ NADP+ FAD CoA リポ酸 TPP PALP THF UDPG CoQ Biotin Coenzyme B12 S-adenosy.l-methionine |
 |
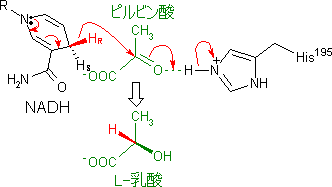 乳酸デヒドロゲナーゼのNADHから,Hがピルビン酸に移動する。同時に,酵素の活性中心のHis-195のイミダゾール基のHがピルビン酸のカルボニル基のOによって引き抜かれてOHとなり,乳酸が生じる。この反応は立体選択的で,生じる乳酸はL型である。 |
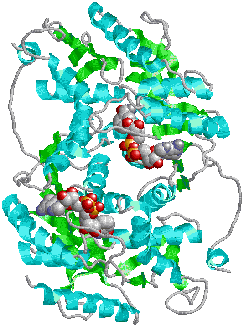
| ブタ筋肉乳酸デヒドロゲナーゼ(二量体) 中央に2分子のNADHが結合 |
乳酸デヒドロゲナーゼの触媒機構 |
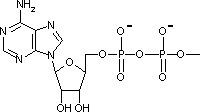
 アデノシン三リン酸 (Adenosine triphosphate, ATP)
アデノシン三リン酸 (Adenosine triphosphate, ATP)
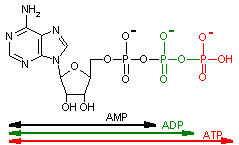 |
ATPは次のようにしてつくられる。 |
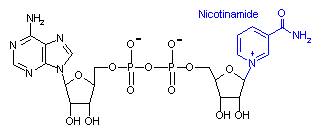
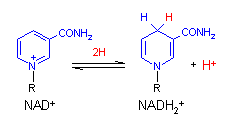
ニコチンアミドアデニンジヌクレオチド (Nicotinamide adenine dinucleotide, NAD+)
 |
 |
ニコチンアミドアデニンジヌクレオチドリン酸 (Nicotinamide adenine dinucleotide phosphate, NADP+)
 |
酸化還元反応において水素の授受に関与する。構造的にはNAD+とはリン酸基が付いただけの違いで,ともにデヒドロゲナーゼの補酵素となる。しかし、NAD+とは異なり、異化代謝ではなく、通常,脂肪酸合成や光合成のような同化代謝で利用される。 |
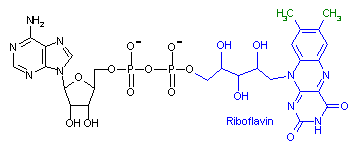
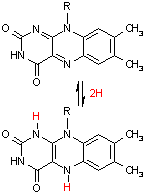
フラビンアデニンジヌクレオチド (Flavin adenine dinucleotide, FAD)
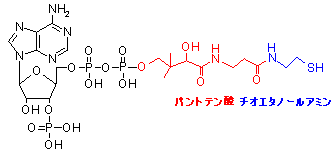
 補酵素A (Coenzyme A, CoA)
補酵素A (Coenzyme A, CoA)
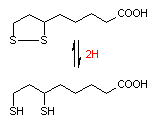
 リポ酸 (a-Lipoic acid)
リポ酸 (a-Lipoic acid)
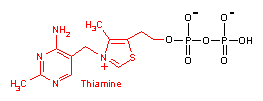
 チアミンピロリン酸 (Thiamine pyrophosphate, TPP)
チアミンピロリン酸 (Thiamine pyrophosphate, TPP)
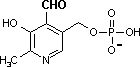
 ピリドキサルリン酸 (Pyridoxal phosphate, PALP)
ピリドキサルリン酸 (Pyridoxal phosphate, PALP)
 テトラヒドロ葉酸 (Tetrahydrofolic acid, THF, Coenzyme F)
テトラヒドロ葉酸 (Tetrahydrofolic acid, THF, Coenzyme F)
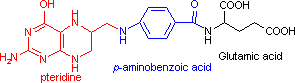
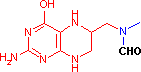 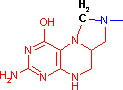 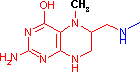 N10-ホルミル-THF N5,N10-メチレン-THF N5-メチル-THF |
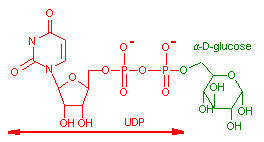
UDPグルコース (Uridine diphosphate glucose, UDPG)
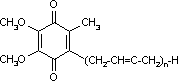
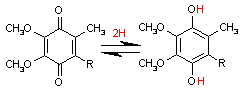
 補酵素Q (Coenzyme Q, CoQ)
補酵素Q (Coenzyme Q, CoQ)
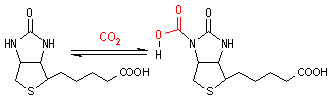
 ビオチン (Biotin, Coenzyme R)
ビオチン (Biotin, Coenzyme R)
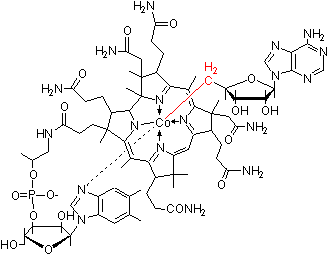
 補酵素B12 (Cobamide, Coenzyme B12)
補酵素B12 (Cobamide, Coenzyme B12)
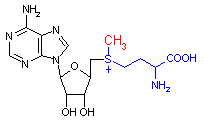
 S-アデノシルメチオニン (S-Adenosyl-L-methionine)
S-アデノシルメチオニン (S-Adenosyl-L-methionine)
|
|